

| Cardiology Research, ISSN 1923-2829 print, 1923-2837 online, Open Access |
| Article copyright, the authors; Journal compilation copyright, Cardiol Res and Elmer Press Inc |
| Journal website https://www.cardiologyres.org |
Original Article
Volume 14, Number 6, December 2023, pages 453-463

Inhibition of the STIM1/Orai1 Signaling Pathway by Glycine Betaine Mitigates Myocardial Hypertrophy in Spontaneous Hypertension Rats
Bing Xiaoa, b, Yan Zhaoa, b, Ke Ke Wanga, Xiu Chun Yanga, Hai Juan Hua, Yue Lia, Yun Fei Xua, Zhen Tian Zhanga, Shuai Wanga, Jing Chao Lua, c
aDepartment of Cardiology, The Second Hospital of Hebei Medical University, Shijiazhuang, China
bThese authors contributed equally to this work.
cCorresponding Author: Jing Chao Lu, Department of Cardiology, The Second Hospital of Hebei Medical University, Shijiazhuang 050000, China
Manuscript submitted September 26, 2023, accepted December 7, 2023, published online December 29, 2023
Short title: GB Alleviating Myocardial Hypertrophy in SHRs
doi: https://doi.org/10.14740/cr1583
| Abstract | ▴Top |
Background: Spontaneous hypertension is a leading risk factor for cardiovascular diseases morbidity and mortality. Glycine betaine (GB) is a natural vitamin that has the potential to lower blood pressure. This work attempted to investigate the role and mechanisms of GB in spontaneous hypertension.
Methods: Spontaneously hypertensive rats (SHRs) were administrated with 100, 200, or 400 mg/kg of GB by gavage or combined with by injection of lentivirus-mediated STIM1 overexpression vector. The heart rate (HR), systolic blood pressure (SBP), diastolic blood pressure (DBP) and heart weight/body weight (HW/BW) of rats were monitored. The pathological changes in myocardium were examined by hematoxylin and eosin staining and Masson staining. The expression of genes and proteins was detected by quantitative real-time PCR, western blotting, and immunohistochemistry.
Results: GB at 200 and 400 mg/kg reduced the HR, SBP, DBP and HW/BW in SHRs. GB decreased the cross-sectional area and fibrotic area in the myocardium and downregulated the expression of atrial natriuretic peptide (ANP) and β-myosin heavy chain (β-MHC) in the myocardium of SHRs. It indicated that GB treatment effectively alleviated myocardial hypertrophy in SHRs. Additionally, GB treatment repressed the expression of stromal interaction molecule 1 (STIM1) and calcium release-activated calcium channel protein 1 (Orai1) in the myocardium of SHRs. STIM1 overexpression reversed GB treatment-mediated inhibition of myocardial hypertrophy in SHRs.
Conclusions: In conclusion, GB repressed STIM1/Orai1 signaling pathway, which contributed to alleviating myocardial hypertrophy in SHRs. Thus, our study provides a theoretical basis for GB as an antihypertensive drug.
Keywords: Glycine betaine; STIM1; Orai1; Myocardial hypertrophy; Spontaneous hypertension
| Introduction | ▴Top |
Primary hypertension, also known as spontaneous hypertension, is a cardiovascular syndrome with elevated systemic arterial pressure as the main clinical manifestation. Poor long-term blood pressure control will lead to corresponding changes in the heart structure, that is, left ventricular hypertrophy [1]. Left ventricular hypertrophy increases the risk of myocardial ischemia, sudden cardiac death, heart failure, arrhythmia, and other cardiovascular diseases in hypertensive patients [2]. Hypertension affects more than 1.2 billion individuals worldwide, and the incidence of hypertension is increasing year by year [3]. As a result, spontaneous hypertension has become the most critical and expensive public health problem. Therefore, it is of great significance to find therapeutic targets and drugs that can delay or even reverse left ventricular hypertrophy for the treatment of spontaneous hypertension.
Glycine betaine (GB; N, N, N-trimethyl amine) is a natural vitamin that exists widely in animals and plants. GB has dual physiological functions of regulating osmotic pressure and methyl donor [4]. As an osmotic buffer, GB maintains the normal osmotic pressure of cells, inhibits the denaturation of proteins and enzymes, thus protecting cells from osmotic damage caused by environmental stress [5]. In addition, GB plays a biological role as a methyl donor in the methionine cycle biochemical reaction [6]. Under the action of GB and homocysteine methyltransferase, GB decreases the levels of plasma homocysteine by promoting the process of homocysteine methylation, thus reducing the risk factors of cardiovascular diseases, including hypertension [7, 8].
Stromal interaction molecule 1 (STIM1) is mainly distributed on the endoplasmic reticulum (ER) membrane, which is a calcium ion sensor for store-operated calcium channels [9]. Calcium release-activated calcium channel protein 1 (Orai1) is a calcium channel localized on the cytoplasmic membrane, which mediates short-term calcium homeostasis and long-term calcium signaling [10]. STIM1 and Orai1 work together to maintain intracellular and extracellular Ca2+ balance [11]. STIM1 and Orai1 function can be modulated by a wide range of different types of post-translational modification, including phosphorylation, glycosylation, and oxidation by reactive oxygen species (ROS) [12-14]. A previous study has confirmed that silencing of STIM1 in smooth muscle attenuates angiotensin II-induced pathological features such as hypertension and myocardial hypertrophy in mice [15]. STIM1 promotes vascular smooth muscle contraction and store-operated calcium entry by physically and functionally interacting with tropomyosin 3, and thus contributes to high-salt intake-induced hypertension in rats [16]. All these findings suggest that STIM1/Orai1 is involved in the pathological progression of spontaneous hypertension. In this study, we speculated that GB could improve hypertension-induced myocardial hypertrophy by regulating STIM1/Orai1 signaling pathway.
| Materials and Methods | ▴Top |
Animals
Male Wistar-Kyoto (WKY) rats and spontaneously hypertensive rats (SHRs) (7 - 8 weeks old, weighting 200 - 230 g) were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd. (China). Rats were housed under (specific pathogen free) SPF conditions with a constant temperature (20 - 24 °C) and humidity (40-60%). After 4 weeks of adaptation, the experiment protocol started when rats were at 11 - 12 weeks.
All protocols were approved by the Ethics Committee of the Second Hospital of Hebei Medical University (No. 2022-AE154). Animal experiments were followed the Guidelines for the Ethical Review of Laboratory Animal Welfare (China National Standard GB/T 35892-2018).
Animal groups
Rats were randomly divided into seven groups that had eight rats per group. GB (98% purity, MCE, Shanghai, China) was freshly prepared in 0.1 M citrate buffer, pH 4.5. The vector pcDNA3.1 carrying full length of STIM1 (pcDNA3.1-STIM1) and negative control vector (pcDNA3.1-NC) were packaged into lentivirus (LV) particles, generating LV-STIM1 and LV-NC (GenePharma, Shanghai, China). SHR rats were injected with LV-STIM1 or LV-NC into the caudal vein at 2 × 1010 transducing units (TU)/dose per rat. Treatment started as follows: 1) Control group treated with phosphate buffered saline (PBS) (WKY) (n = 8): WKY rats received PBS via oral gavage daily for 21 days; 2) Hypertension group treated with PBS (SHR) (n = 8): SHR rats received PBS via oral gavage daily for 21 days; 3) Hypertension group treated with 100 mg/kg/day GB (SHR + 100 mg/kg/day GB) (n = 8): SHR rats received 100 mg/kg GB via oral gavage daily for 21 days; 4) Hypertension group treated with 200 mg/kg/day GB (SHR + 200 mg/kg/day GB) (n = 8): SHR rats received 200 mg/kg GB via oral gavage daily for 21 days; 5) Hypertension group treated with 400 mg/kg/day GB (SHR + 400 mg/kg/day GB) (n = 8): SHR rats received 400 mg/kg GB via oral gavage daily for 21 days; 6) Hypertension group treated with 400 mg/kg/day GB and LV-NC (GB + LV-NC) (n = 8): SHR rats was injected with LV-NC into the caudal vein at the first day and received 400 mg/kg GB via oral gavage daily for 21 days; 7) Hypertension group treated with 400 mg/kg/day GB and LV-STIM1 (GB + LV-STIM1) (n = 8): SHR rats was injected with LV-STIM1 into the caudal vein at the first day to overexpress STIM1 and received 400 mg/kg GB via oral gavage daily for 21 days.
The treatment protocol of the groups and the methods were shown here (Supplementary Material 1, www. cardiologyres.org). During the modeling process, the heart rate (HR), systolic blood pressure (SBP) and diastolic blood pressure (DBP) in rats were monitored every 2 days utilizing the noninvasive tail cuff method (CODA Eight-Channel High Throughput Non-Invasive Blood Pressure System, Kent Scientific, Boston, MA, USA) as previous reported [17]. At the end of the 3-week period (21 days), the rats were euthanized with an overdose of pentobarbital sodium (200 mg/kg). Heart tissues were separated from the euthanized rats. The heart weight (HW) and body weight (BW) were weighted to calculate the ratio of HW/BW. The left ventricular myocardium was collected for histological analysis, quantitative real-time PCR (qRT-PCR), western blotting and immunohistochemistry (IHC) staining.
Hematoxylin and eosin (H&E) staining
The heart and left ventricular myocardia tissues were fixed in 4% paraformaldehyde for 24 h, and then embedded in paraffin. Following dewaxing and hydration, serial myocardium sections with 4 µm were collected. Sections of heart tissues were stained with hematoxylin for 5 min and then stained with eosin for 30 s applying H&E Staining Kit (Beyotime, Shanghai, China). Sections of left ventricular myocardia tissues were stained with Modified Masson’s Trichrome Stain Kit (Solarbio, Beijing, China).
Wheat germ agglutinin (WGA) staining
Following dewaxing and hydration, serial myocardium sections with 4 µm were collected. After sections of heart tissues are slightly dry, draw a circle on the objective tissue with a liquid blocking pen. Cover the marked area with diluted WGA solution (W21405, Thermo Fisher Scientific, Waltham, MA, USA) and incubate in a dark place at 37 °C for 30 min. The rocker unit was washed three times with PBS (pH 7.4) for 5 min each time, and then incubated with 4’,6-diamidino-2-phenylindole (DAPI) solution at room temperature for 10 min for nuclear re-staining and storage away from light. Wash three times with PBS (pH 7.4) in a Rocker device, 5 min each. Throw away liquid slightly, then coverslip with anti-fade mounting medium. The sections were observed under a fluorescent microscope, and analyzed using Image J software (NIH, Bethesda, MD, USA). Five fields were randomly selected from each myocardial tissue section for cross-sectional area statistics, and each field had at least 50 cells. After calculating the average cross-sectional area of cardiomyocyte in each field, the average value of the five fields was taken as the final result.
IHC staining
Paraffin sections of left ventricular myocardium tissues were treated with 3% H2O2 for 10 min, followed by antigen retrieval with citrate buffer. Nonspecific sites were blocked with bovine serum albumin (BSA; Meilunbio, Dalian, China). Sections were incubated with anti-STIM1 (1:100; MA1-19451, Thermo Fisher Scientific, Waltham, MA, USA) and then treated with biotin-conjugated immunoglobulin G (IgG) antibody (1:1,000; PA1-29634, Thermo Fisher Scientific, Waltham, MA, USA). Sections were stained with diaminobenzidine and then counterstained with hematoxylin. The sections were observed under an optical microscope. To evaluate fibrosis, areas of interest (perivascular, and interstitial areas) were defined in Image J. Five fields were randomly selected, and the average value was used as the value of fibrosis area. The extent of fibrosis was determined by calculating the area of perivascular fibrosis, the sum of the interstitial fibrosis areas as a percentage of the total area of the entire section.
Plasma homocysteine measurement
During blood collection, the anesthetized rats were supine fixed on the fixation plate, the hair in the anterior cardiac area was cut off, the skin was disinfected with iodine alcohol, the chest was opened, the heart was exposed, and the right ventricle was pierced with a needle to suck blood. After the blood is removed from the heart, it is centrifuged at 3,500 rpm for 15min, the supernatant is absorbed as plasma. Plasma homocysteine concentration was determined using the Homocysteine Colorimetric Assay Kit (E-BC-K854-M, ElabScience, Waltham, MA, USA) by spectrophotometry using an enzyme-linked immunosorbent assay (ELISA) reader (Multiskan FS, Thermo Fisher Scientific, Waltham, MA, USA).
qRT-PCR
Total RNA was extracted from the left ventricular myocardium applying TRIzol reagent (Invitrogen). RNA integrity was examined by performing 1.5% agarose gel electrophoresis. Total RNA was served as template and was reversed transcribed to cDNA using PrimeScript™ RT reagent Kit (Takara, Tokyo, Japan). TB Green® Premix Ex Taq™ II (Takara) was used to detect the relative expression of genes. PCR reaction mixture (25 µL) contained 12.5 µL TB Green® Premix Ex Taq™, 0.5 µL forward primer, 0.5 µL reverse primer, 2 µL cDNA and 9.5 µL sterile water. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) served as internal control. The primer sequences were listed in Table 1. The results were analyzed using 2-ΔΔCT method for quantification.
 Click to view | Table 1. Primers Used in qRT-PCR |
Western blotting
Total protein was extracted from the left ventricular myocardium utilizing Total Protein Extraction Kit (Solarbio). Protein samples were separated by 10% sodium dodecyl-sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gel electrophoresis and transferred onto the nitrocellulose membranes. The membranes were incubated with the primary antibodies, anti-atrial natriuretic peptide (ANP) (1:1,000 dilution; PA5-29559, Thermo Fisher Scientific, Waltham, MA, USA), anti-β-myosin heavy chain (β-MHC) (1:1,000 dilution; ab37484, Abcam, Cambridge, MA, USA), anti-STIM1 (1:1,000 dilution; MA1-19451, Thermo Fisher Scientific, Waltham, MA, USA), anti-Orai1 (1:1,000 dilution; ab111960, Abcam, Cambridge, MA, USA) at 4 °C overnight after blocking with 5% skimmed milk. The membranes were stained with horseradish peroxidase (HRP)-conjugated anti-rabbit IgG antibody (1:5,000; ab270144, Abcam, Cambridge, MA, USA) or HRP-conjugated anti-rat IgG antibody (1:5,000; ab7097, Abcam, Cambridge, MA, USA) at room temperature. GAPDH antibody (1:5,000; ab8245, Abcam, Cambridge, MA, USA) served as internal control. The bands were developed by enhanced chemiluminescence reagent (Yeasen, Shanghai, China), and the data were analyzed by Image J software. The resulting ratios, expressed as fold change, were used to compare relative protein levels across the samples on blots.
Statistical analysis
Each assay was performed three times. Data were expressed as mean ± standard deviation (SD). Statistical analysis was carried out applying SPSS 22.0 statistical software (IBM, Armonk, NY, USA). Two-tailed Student’s t-test, one-way analysis of variance (ANOVA) and two-way ANOVA was used to analyze the statistical difference. P value less than 0.05 was considered as a significant difference.
| Results | ▴Top |
GB alleviated myocardial hypertrophy in SHRs
To determine the biological role of GB in spontaneous hypertension, SHRs were administrated with different dosages of GB. The monitoring results of HR, SBP and DBP revealed that SHRs exhibited higher levels of HR, SBP and DBP as compared with WKY rats. GB at 100 mg/kg/day slightly reduced HR, SBP and DBP in SHRs, while GB at 200 and 400 mg/kg/day significantly repressed the HR, SBP and DBP in SHRs (Fig. 1a-c). The ratio of HW/BW increased in SHRs. GB at 200 and 400 mg/kg/day effectively reduced the HW/BW ratio in SHRs (Fig. 1d).
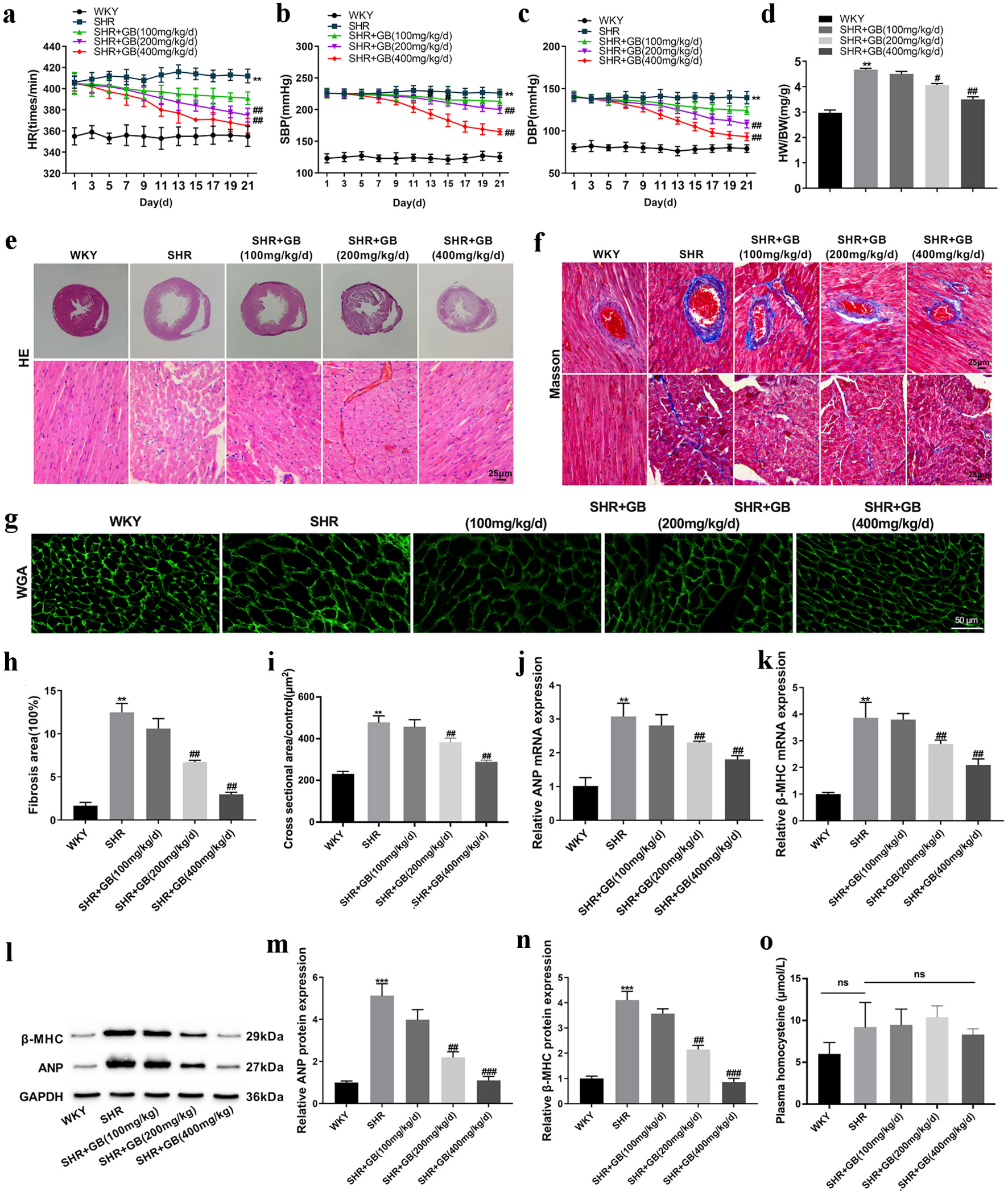 Click for large image | Figure 1. The effect of GB on myocardial hypertrophy of SHRs. SHRs were administrated with 100, 200 or 400 mg/kg/day of GB. Normal WKY rats served as control. Each group included n = 8 rats. The HR (a), SBP (b) and DBP (c) of rats were monitored. (d) The HW/BW of rats was detected at 21 days. (e) Images of hematoxylin and eosin (H&E) staining of the myocardial tissue. (g, i) WGA staining examined the myocyte cross-sectional area of rats. (f, h) Masson staining detected fibrosis in the myocardium of rats. (j-k) The qRT-PCR analyzed the expression of ANP (j) and β-MHC (k) in the myocardium of rats. (l-n) Western blotting detected the expression of ANP (m) and β-MHC (n) in the myocardium of rats. The representative blot was shown from three independent repeats. (o) The effect of GB on plasma homocysteine of SHRs (n = 5). The values are presented as mean ± SD. **P < 0.01, ***P < 0.001 vs. WKY group; #P < 0.05, ##P < 0.01, ###P < 0.001 vs. SHR group. GB: glycine betaine (N, N, N-trimethyl glycine); SHRs: spontaneously hypertensive rats; WKY: Wistar-Kyoto; WGA: wheat germ agglutinin; BW: body weight; DBP: diastolic blood pressure; HR: heart rate; HW: heart weight; qRT-PCR: quantitative real-time PCR; SBP: systolic blood pressure; ANP: atrial natriuretic peptide; β-MHC: β-myosin heavy chain; SD: standard deviation. |
Additionally, H&E staining examined the impact of GB on myocardial hypertrophy in SHRs. WKY rats exhibited orderly arranged myocardial fibers and myocardial cells with normal psychological structure. In SHR rats, the arrangement of myocardial fibers was disordered, the myocardial cell bundle was broken, the myocardial cells were significantly enlarged, and the cross-sectional area of myocardial cells was significantly increased. These pathological changes were effectively alleviated by GB treatment (200 and 400 mg/kg/day) (Fig. 1e, g, h). The results of Masson staining revealed that fibrosis area was elevated in the myocardium of SHRs. The fibrosis area in the myocardium of SHRs was severely inhibited by GB treatment (200 and 400 mg/kg/day) (Fig. 1f, i).
Subsequent analysis of the mRNA and protein expression of hypertrophic genes (ANP and β-MHC) showed similar trends (Fig. 1j-n). All these data indicated that GB treatment alleviated myocardial hypertrophy in SHRs. Plasma homocysteine concentration was measured by the commercial ELISA kit. No significant difference was observed between the groups in plasma homocysteine concentration (Fig. 1o).
GB treatment repressed STIM1/Orai1 signaling pathway in SHRs
The influence of GB on STIM1 and Orai1 expression in SHRs was detected. The results of qRT-PCR showed that the expression of STIM1 and Orai1 was elevated in the myocardium of SHRs. GB treatment (200 and 400 mg/kg/day) caused a downregulation of STIM1 and Orai1 in the myocardium of SHRs (Fig. 2a, b). Higher expression of STIM1 and Orai1 was observed in the myocardium of SHRs, which was repressed by GB treatment (200 and 400 mg/kg/day) (Fig. 2c-e). Applying IHC staining, GB treatment (200 and 400 mg/kg/day) reduced the expression of STIM1 in the myocardium of SHRs (Fig. 2f). Thus, GB repressed STIM1/Orai1 signaling pathway in SHRs.
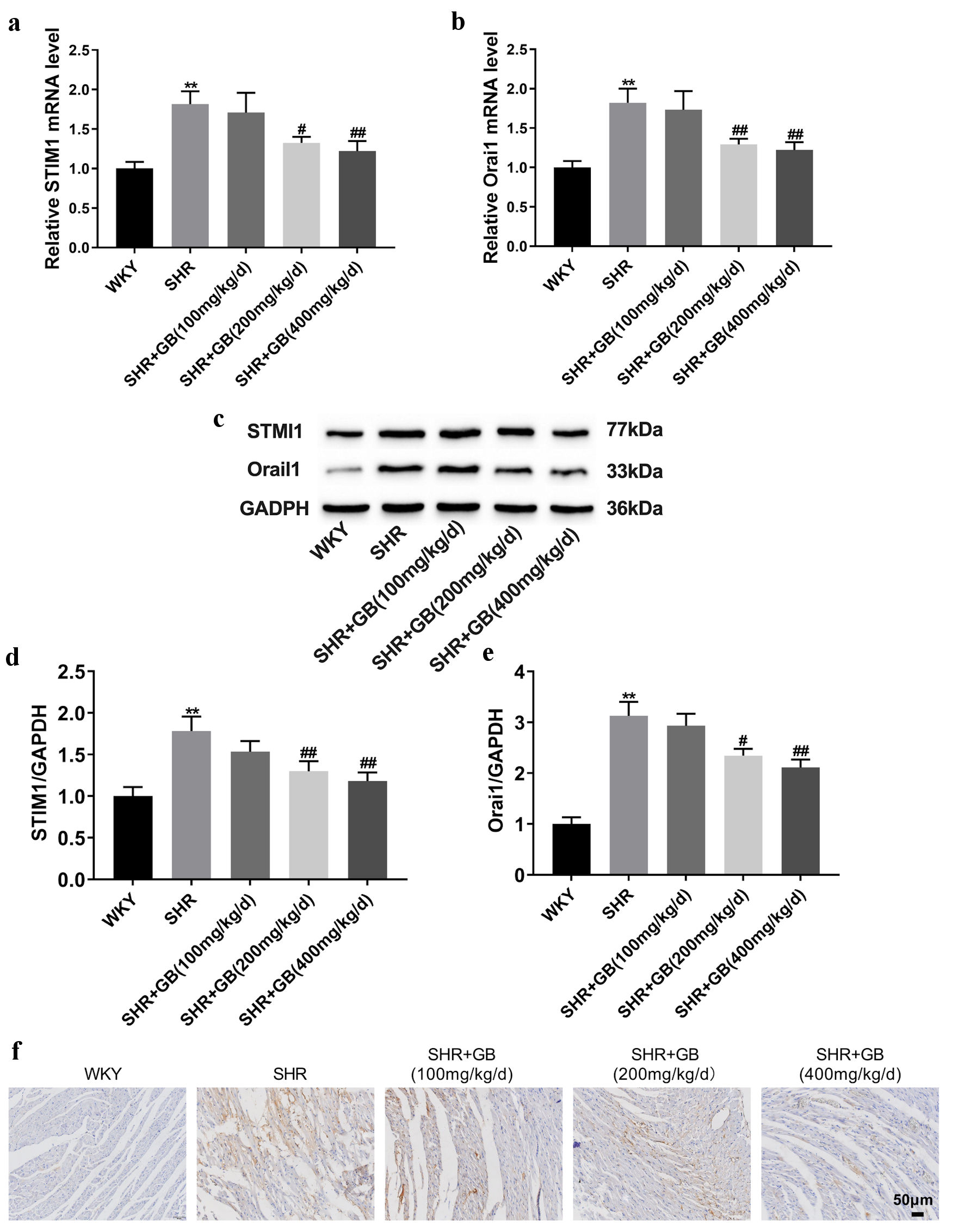 Click for large image | Figure 2. The impact of GB on the expression of STIM1 and Orai1 in SHRs. SHRs were administrated with 100, 200 or 400 mg/kg/day of GB. Normal WKY rats served as control. (a, b) The qRT-PCR analyzed the expression of STIM1 (a) and Orai1 (b) in the myocardium of rats. (c-e) Western blotting analyzed the expression of STIM1(d) and Orai1 (e) in the myocardium of rats. The representative blot was shown from three independent repeats. (f) IHC staining examined the expression of STIM1 in the myocardium of rats. Each group included n = 8 rats. The values are presented as mean ± SD. **P < 0.01 vs. WKY group; #P < 0.05, ##P < 0.01 vs. SHR group. GB: glycine betaine (N, N, N-trimethyl glycine); IHC: immunohistochemistry; Orai1: calcium release-activated calcium channel protein 1; qRT-PCR: quantitative real-time PCR; SHRs: spontaneously hypertensive rats; STIM1: stromal interaction molecule 1; WKY: Wistar-Kyoto; SD: standard deviation; GAPDH: glyceraldehyde-3-phosphate dehydrogenase. |
STIM1 overexpression reversed the impact of GB on myocardial hypertrophy in SHRs
Next, we investigated the molecular mechanism of GB in regulating myocardial hypertrophy by overexpressing STIM1 in vivo. SHRs were injected with LV-STIM1 or LV-NC into the caudal vein and administrated with 400 mg/kg/day of GB for 21 days by gavage. GB treatment notably reduced the levels of HR, SBP, and DBP in SHRs. The influence conferred by GB treatment was significantly abolished by STIM1 overexpression (Fig. 3a-c). Moreover, STIM1 overexpression impaired the GB-mediated decrease of HW/BW ratio in SHRs (Fig. 3d). The results obtained from H&E, WGA and Masson staining uncovered that GB treatment inhibited the myocyte cross-sectional area and fibrosis area in SHRs, which was effectively abrogated by upregulated STIM1 (Fig. 4a-e). Taken together, STIM1 overexpression reversed the impact of GB on myocardial hypertrophy in SHRs.
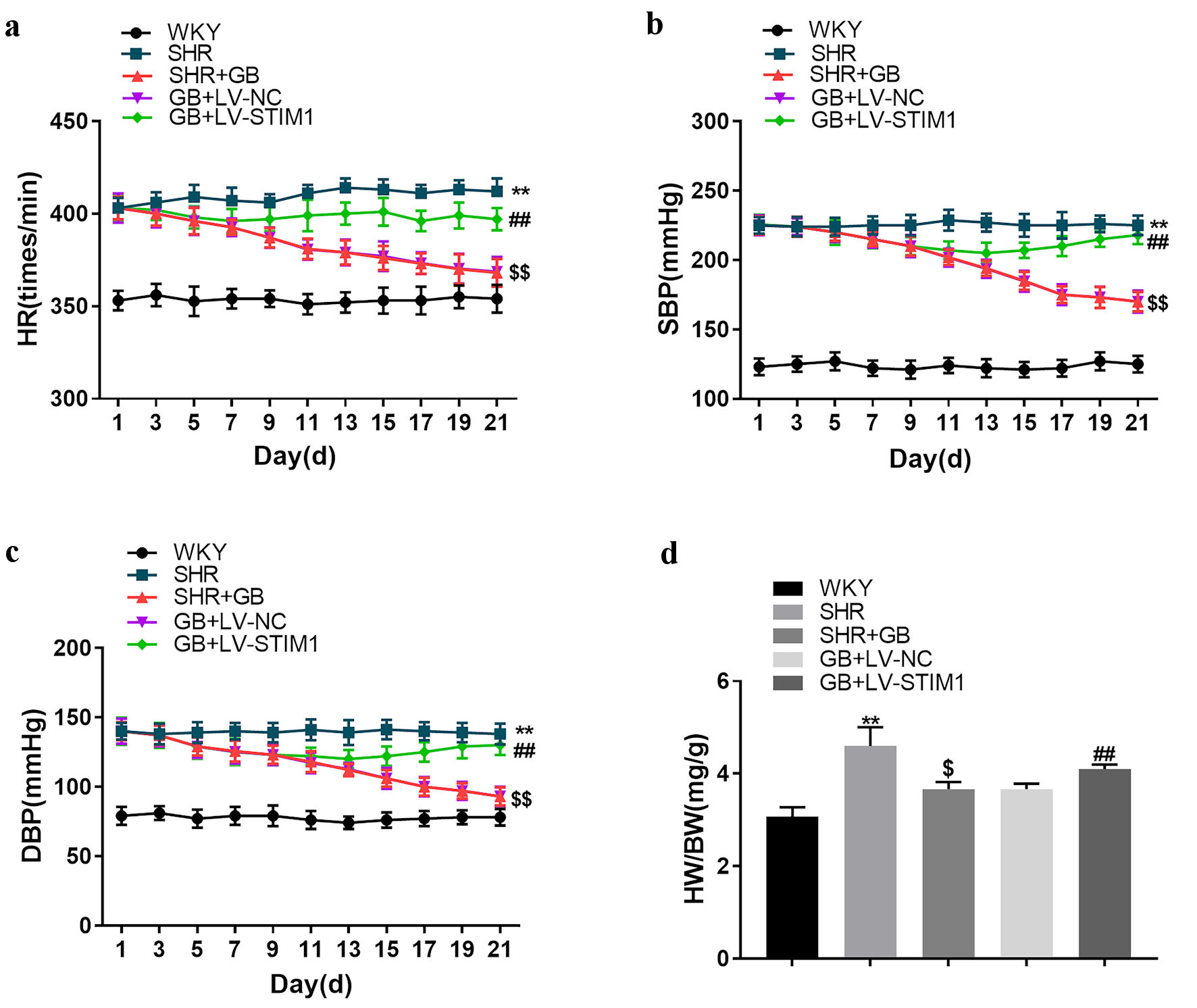 Click for large image | Figure 3. The impact of STIM1 overexpression on SHRs. SHRs were administrated with 400 mg/kg/day of GB or followed by injection of LV-STIM1 or LV-NC. Normal WKY rats served as control. The HR (a), SBP (b) and DBP (c) of rats were monitored. (d) The HW/BW of rats was detected. Each group included n = 8 rats. The values are presented as mean ± SD. **P < 0.01 vs. WKY group; $P < 0.05 vs. $$P < 0.01 vs. SHR group; ##P < 0.01 vs. GB + LV-NC group. BW: body weight; DBP: diastolic blood pressure; GB: glycine betaine (N, N, N-trimethyl glycine); HR: heart rate; HW: heart weight; LV: lentivirus; NC: negative control; SBP: systolic blood pressure; SHRs: spontaneously hypertensive rats; STIM1: stromal interaction molecule 1; WKY: Wistar-Kyoto; SD: standard deviation. |
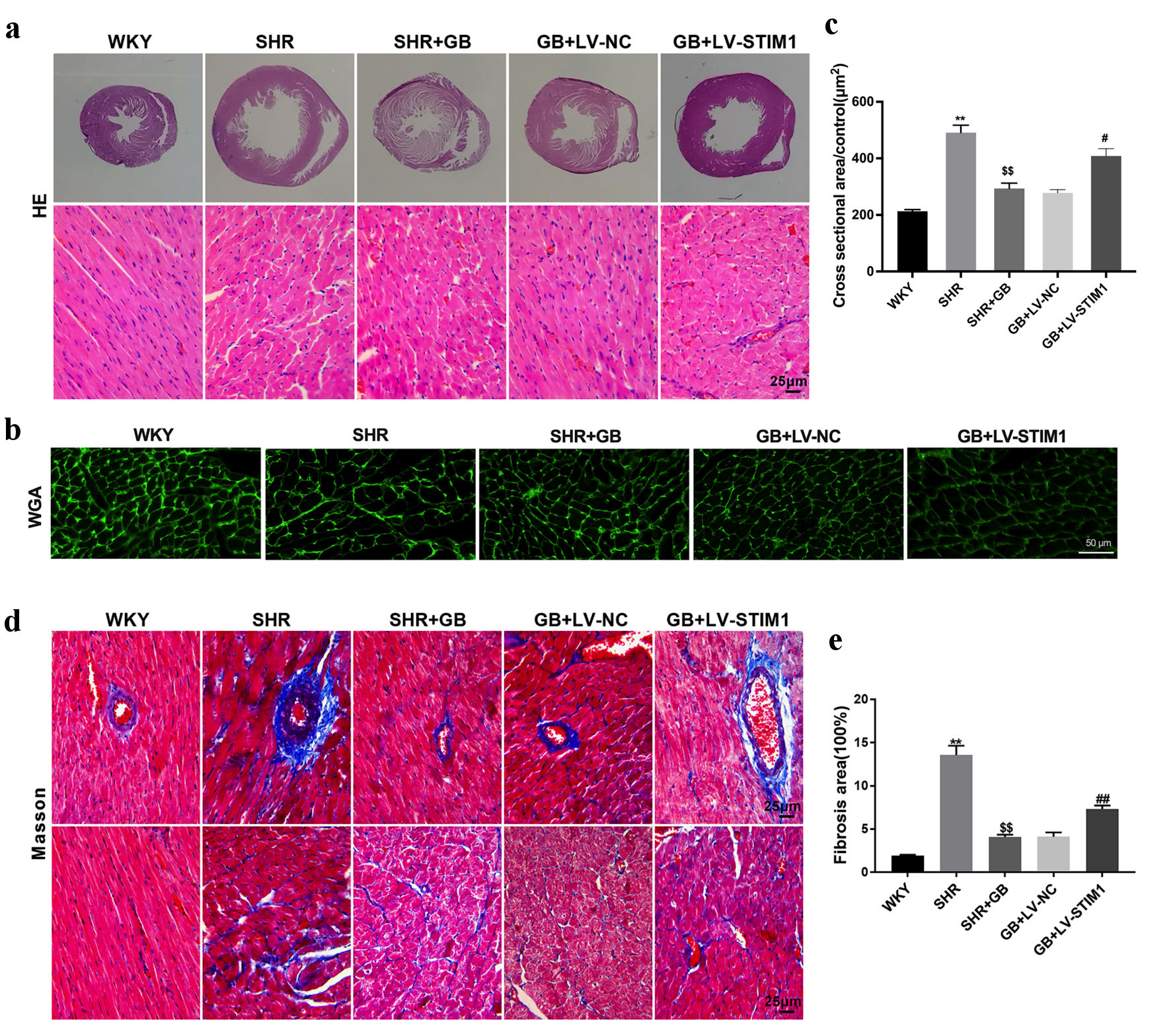 Click for large image | Figure 4. The impact of STIM1 overexpression on myocardial hypertrophy of SHRs. SHRs were administrated with 400 mg/kg/day of GB or followed by injection of LV-STIM1 or LV-NC. Normal WKY rats served as control. (a) Images of hematoxylin and eosin (H&E) staining of the myocardial tissue. (b, c) WGA staining examined the myocyte cross-sectional area of rats. (d, e) Masson staining detected cardiac fibrosis of rats. Each group included n = 8 rats. The values are presented as mean ± SD. **P < 0.01 vs. WKY group; $$P < 0.01 vs. SHR group; #P < 0.05, ##P < 0.01 vs. GB + LV-NC group. GB: glycine betaine (N, N, N-trimethyl glycine); LV: lentivirus; NC: negative control; SHRs: spontaneously hypertensive rats; STIM1: stromal interaction molecule 1; WKY: Wistar-Kyoto; WGA: wheat germ agglutinin; SD: standard deviation. |
GB alleviated myocardial hypertrophy in SHRs by regulating STIM1/Orai1 signaling pathway
To further verify the role of STIM1/ Orai1 in myocardial hypertrophy, we examined the level of ANP, β-MHC following STIM1 overexpression. As expected, GB treatment suppressed the expression of ANP, β-MHC, STIM1 and Orai1 in SHRs. STIM1 overexpression reversed GB-mediated downregulation of ANP, β-MHC, STIM1 and Orai1 in SHRs (Fig. 5a-e). Results of IHC staining showed that GB-mediated decreased expression of STIM1 was abrogated by LV-STIM1 (Fig. 5f). Thus, GB alleviated myocardial hypertrophy in SHRs by regulating STIM1/Orai1 signaling pathway.
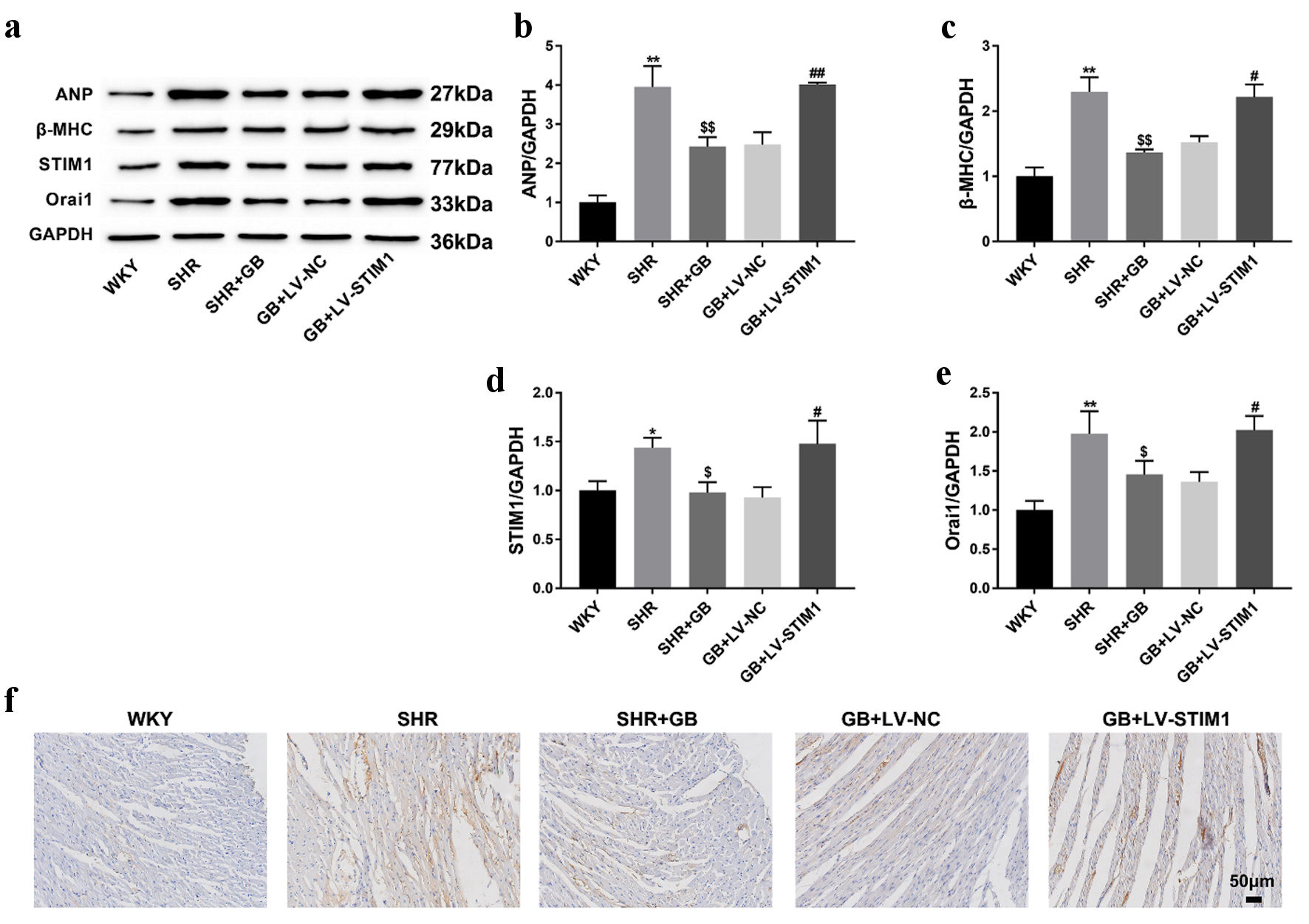 Click for large image | Figure 5. The impact of GB treatment on the expression of STIM1 and Orai1 in SHRs following overexpressed STIM1. SHRs were administrated with 400 mg/kg/day of GB or followed by injection of LV-STIM1 or LV-NC. Normal WKY rats served as control. (a-e) Western blotting analyzed the expression of ANP (b), β-MHC (c), STIM1 (d) and Orai1 (e) in the myocardium of rats. The representative blot was shown from three independent repeats. (f) IHC staining examined the expression of STIM1 in the myocardium of rats. Each group included n = 8 rats. The values are presented as mean ± SD. *P < 0.05, **P < 0.01 vs. WKY group; $P < 0.05, $$P < 0.01 vs. SHR group; #P < 0.05, ##P < 0.01 vs. GB + LV-NC group. GB: glycine betaine (N, N, N-trimethyl glycine); LV: lentivirus; NC: negative control; SHRs: spontaneously hypertensive rats; STIM1: stromal interaction molecule 1; WKY: Wistar-Kyoto; SD: standard deviation; ANP: atrial natriuretic peptide; β-MHC: β-myosin heavy chain; IHC: immunohistochemistry; Orai1: calcium release-activated calcium channel protein 1; GAPDH: glyceraldehyde-3-phosphate dehydrogenase. |
| Discussion | ▴Top |
In this study, we investigated the role and mechanism of GB in spontaneous hypertension. Our data showed that GB treatment alleviated SBP, DBP, HW/BW, and myocardial hypertrophy in a concentration-dependent manner. GB treatment repressed the expression of STIM1 and Orai1 in the myocardium of SHRs. Conversely, STIM1 overexpression reversed GB induced SBP, DBP, HW/BW, and myocardial hypertrophy. Therefore, our study demonstrates that GB alleviates myocardial hypertrophy in SHRs by regulating STIM1/Orai1 signaling pathway.
GB (N, N, N-trimethyl glycine) is a quaternary amine isolated for the first time from beets [7]. Initially, GB was studied as an osmolyte accumulated in the cells and the key to regulating cell volume and swelling [18]. Later, GB was demonstrated as the main methyl donor in the methionine cycle and was widely used in the treatment of hyperhomocysteinemia to prevent the development of coronary atherosclerosis, cerebrovascular disease, and peripheral vascular disease [7, 19-22]. GB ameliorated sulfur amino acid (SAA) metabolism, which was involved in essential metabolic pathways like glutathione synthesis, to protect against oxidative stress. Additionally, several studies indicated that GB suppressed the NF-κB, NLRP3 and TLR4 pathways, leading to a decrease in inflammatory cytokine [23-25]. Betaine also inhibited caspase-3 to reduce apoptosis and repair ER stress [26]. Another study showed that betaine pretreatment protected against photochemically-induced thrombosis in pial arterioles and venules in vivo and platelet aggregation in vitro. They also observed that betaine pretreatment decreased lipid peroxidation in plasma [27].
The functional role of GB in hypertension was first reported as early as 1951. Hypertensive subjects experienced transient reduced blood pressure in the combination of betaine and guanidinoacetate [28]. In a large population-based study of 7,074 mid-aged and elderly subjects, plasma betaine was reported to have a negative relationship with blood pressure level [29]. Similar results were observed in dialysis patients that circulating GB was negatively correlated with blood pressure [30]. As for the animal experiments, Rosas-Rodriguez et al reported that GB concentration increased during physiological cardiac hypertrophy induced by pregnancy in Sprague-Dawley rats, which was regulated by the enzyme betaine aldehyde dehydrogenase (BADH) to protect myocytes from hyperosmotic stress [31]. In the current study, we observed that GB reduced both SBP and DBP, and alleviated myocardial hypertrophy in SHRs. Our findings indicated that GB may be a potential antihypertensive drug.
The influence of betaine supplementation on circulating homocysteine concentration has been studied mainly in clinical settings [32, 33]. Betaine supplementation has an effective agent for decreasing plasma homocysteine in healthy adults. Specifically, the daily ingestion of 6 g betaine for 6 weeks decreased fasting plasma homocysteine by about 11% [32]. A meta-analysis which included 12 studies examined the overall effect of betaine supplementation on homocysteine concentrations, and the results indicated that betaine supplementation had homocysteine-lowering effects. The minimum dosage of betaine evaluated in the study was 1.5 g/day for 6 weeks [34]. In our study, no significant downregulation of plasma homocysteine was observed following GB supplementation, which may be attributed to the short evaluation time and the dosage of GB. Additionally, betaine supplementation was reported to have adverse effects on lipid profiles. Betaine supplementation greater than 4 g/day had a significant effect on concentrations of total cholesterol (TC) and low-density lipoprotein (LDL) [34]. It is well established that improving lipid profile, one of the major contributors to atherosclerosis, can reduce cardiovascular disease (CVD) risk, particularly in people with additional cardiovascular risk factors. Further clinical and experimental studies must be conducted to determine the association between betaine supplementation and its consequence on the lipid profile.
Considerable attention has been focused on abnormalities of Ca2+ cycling in cardiac hypertrophy as a possible therapeutic target [35]. STIM1 and Orai1 are major proteins of store-operated calcium channel, and involved in regulating the progression of various diseases, including hypertension [36, 37]. The expression of STIM1 is increased in angiotensin II-induced hypertension mice, and STIM1 upregulation enhances ER stress to aggravate the development of hypertension [15, 38]. The interaction between STIM1 and tropomyosin 3 contributes to vascular smooth muscle contraction and store-operated calcium entry, thereby accelerating high-salt intake-induced hypertension [16]. However, as far as we are aware, no study has examined the effect of GB on STIM1/Orai1 pathway in myocardial hypertrophy. In the present work, we first reported that GB treatment attenuated myocardial hypertrophy by downregulating STIM1/Orai1 pathway in SHRs. However, since STIM1 and Orai1 are major proteins to regulate intracellular and extracellular Ca2+ flow, whether GB can affect the concentration of intracellular Ca2+ through STIM1/Orai1 signaling pathway in SHRs needs to be confirmed.
There are several limitations in this study. First, the evaluation time is only 21 days in our study. Second, the effects mediated by GB on myocardial hypertrophy may not only via inhibiting inflammation and oxidative stress but also directly via downregulated blood pressure. Whether GB plays a protective role in a blood pressure-dependent manner is not demonstrated in this study. Third, the protective role of GB treatment in myocardial hypertrophy by regulating the STIM1/Orai1 pathway is not performed at the cellular level. And the post-translational modifications of STIM1/Orai1 are not examined. At last, whether GB possesses other biological activities during the pathophysiological process of cardiac hypertrophy, like anti-inflammation and anti-oxidation, needs to be explored in the future.
Conclusions
In conclusion, GB represses STIM1/Orai1 signaling pathway, which contributes to alleviating myocardial hypertrophy in SHRs. Thus, our results shed new light on the mechanism of action of GB in spontaneous hypertension and provide a theoretical basis for GB as an antihypertensive drug.
| Supplementary Material | ▴Top |
Suppl 1. Timeline of animal experiments.
Acknowledgments
None to declare.
Financial Disclosure
This study was supported by the Project of Hebei Natural Science Foundation (No. H2021206205), Key Project of Medical Science Research in Hebei Province (No. 20220102) and Program for Excellent Talents in Clinical Medicine of Hebei Province (No. 303-16-20-09, No. ZF2023148).
Conflict of Interest
None to declare.
Informed Consent
Not applicable.
Author Contributions
All authors contributed to data analysis, drafting, or revising the article, have agreed on the journal to which the article will be submitted, gave final approval of the version to be published, and agreed to be accountable for all aspects of the work.
Data Availability
The data used to support the findings of this study are available from the corresponding author upon reasonable request.
Abbreviations
BADH: betaine aldehyde dehydrogenase; BW: body weight; DBP: diastolic blood pressure; ER: endoplasmic reticulum; GB: glycine betaine (N, N, N-trimethyl glycine); H&E: hematoxylin and eosin; HR: heart rate; HW: heart weight; IHC: immunohistochemistry; LV: lentivirus; Orai1: calcium release-activated calcium channel protein 1; qRT-PCR: quantitative real-time PCR; ROS: reactive oxygen species; SAA: sulfur amino acid; SBP: systolic blood pressure; SHRs: spontaneously hypertensive rats; STIM1: stromal interaction molecule 1; WKY: Wistar-Kyoto; WGA: wheat germ agglutinin
| References | ▴Top |
- Wang Z, Zhang C, Bao H, Huang X, Fan F, Zhao Y, Li J, et al. Value of electrocardiographic left ventricular hypertrophy as a predictor of poor blood pressure control: Evidence from the China stroke primary prevention trial. Medicine (Baltimore). 2018;97(44):e12966.
doi pubmed pmc - Lind L, Sundstrom J. Change in left ventricular geometry over 10 years in the elderly and risk of incident cardiovascular disease. J Hypertens. 2019;37(2):325-330.
doi pubmed - Rossier BC, Bochud M, Devuyst O. The hypertension pandemic: an evolutionary perspective. Physiology (Bethesda). 2017;32(2):112-125.
doi pubmed - Figueroa-Soto CG, Valenzuela-Soto EM. Glycine betaine rather than acting only as an osmolyte also plays a role as regulator in cellular metabolism. Biochimie. 2018;147:89-97.
doi pubmed - Knight LS, Piibe Q, Lambie I, Perkins C, Yancey PH. Betaine in the brain: characterization of betaine uptake, its influence on other osmolytes and its potential role in neuroprotection from osmotic stress. Neurochem Res. 2017;42(12):3490-3503.
doi pubmed - Zou H, Chen N, Shi M, Xian M, Song Y, Liu J. The metabolism and biotechnological application of betaine in microorganism. Appl Microbiol Biotechnol. 2016;100(9):3865-3876.
doi pubmed - Rosas-Rodriguez JA, Valenzuela-Soto EM. The glycine betaine role in neurodegenerative, cardiovascular, hepatic, and renal diseases: Insights into disease and dysfunction networks. Life Sci. 2021;285:119943.
doi pubmed - Arumugam MK, Paal MC, Donohue TM, Jr., Ganesan M, Osna NA, Kharbanda KK. Beneficial effects of betaine: a comprehensive review. Biology (Basel). 2021;10(6):456.
doi pubmed pmc - Gudlur A, Zeraik AE, Hirve N, Hogan PG. STIM calcium sensing and conformational change. J Physiol. 2020;598(9):1695-1705.
doi pubmed pmc - Cai X, Zhou Y, Nwokonko RM, Loktionova NA, Wang X, Xin P, Trebak M, et al. The Orai1 store-operated calcium channel functions as a hexamer. J Biol Chem. 2016;291(50):25764-25775.
doi pubmed pmc - Lunz V, Romanin C, Frischauf I. STIM1 activation of Orai1. Cell Calcium. 2019;77:29-38.
doi pubmed - Johnson J, Blackman R, Gross S, Soboloff J. Control of STIM and Orai function by post-translational modifications. Cell Calcium. 2022;103:102544.
doi pubmed pmc - Dorr K, Kilch T, Kappel S, Alansary D, Schwar G, Niemeyer BA, Peinelt C. Cell type-specific glycosylation of Orai1 modulates store-operated Ca2+ entry. Sci Signal. 2016;9(418):ra25.
doi pubmed - Lopez E, Frischauf I, Jardin I, Derler I, Muik M, Cantonero C, Salido GM, et al. STIM1 phosphorylation at Y(316) modulates its interaction with SARAF and the activation of SOCE and I(CRAC). J Cell Sci. 2019;132(10):jcs226019.
doi pubmed - Kassan M, Ait-Aissa K, Radwan E, Mali V, Haddox S, Gabani M, Zhang W, et al. Essential role of smooth muscle STIM1 in hypertension and cardiovascular dysfunction. Arterioscler Thromb Vasc Biol. 2016;36(9):1900-1909.
doi pubmed pmc - Xu F, Zhu J, Chen Y, He K, Guo J, Bai S, Zhao R, et al. Physical interaction of tropomyosin 3 and STIM1 regulates vascular smooth muscle contractility and contributes to hypertension. Biomed Pharmacother. 2021;134:111126.
doi pubmed - Daugherty A, Rateri D, Hong L, Balakrishnan A. Measuring blood pressure in mice using volume pressure recording, a tail-cuff method. J Vis Exp. 2009;(27):1291.
doi pubmed pmc - Chen TH, Murata N. Glycinebetaine protects plants against abiotic stress: mechanisms and biotechnological applications. Plant Cell Environ. 2011;34(1):1-20.
doi pubmed - Harisa GI. Oxidative stress and paraoxonase activity in experimental selenosis: effects of betaine administration. Biol Trace Elem Res. 2013;152(2):258-266.
doi pubmed - Ganesan B, Buddhan S, Anandan R, Sivakumar R, AnbinEzhilan R. Antioxidant defense of betaine against isoprenaline-induced myocardial infarction in rats. Mol Biol Rep. 2010;37(3):1319-1327.
doi pubmed - Heidari R, Niknahad H, Sadeghi A, Mohammadi H, Ghanbarinejad V, Ommati MM, Hosseini A, et al. Betaine treatment protects liver through regulating mitochondrial function and counteracting oxidative stress in acute and chronic animal models of hepatic injury. Biomed Pharmacother. 2018;103:75-86.
doi pubmed - Lever M, George PM, Slow S, Bellamy D, Young JM, Ho M, McEntyre CJ, et al. Betaine and Trimethylamine-N-Oxide as predictors of cardiovascular outcomes show different patterns in diabetes mellitus: an observational study. PLoS One. 2014;9(12):e114969.
doi pubmed pmc - Go EK, Jung KJ, Kim JM, Lim H, Lim HK, Yu BP, Chung HY. Betaine modulates age-related NF-kappaB by thiol-enhancing action. Biol Pharm Bull. 2007;30(12):2244-2249.
doi pubmed - Lee EK, Jang EJ, Jung KJ, Kim DH, Yu BP, Chung HY. Betaine attenuates lysophosphatidylcholine-mediated adhesion molecules in aged rat aorta: modulation of the nuclear factor-kappaB pathway. Exp Gerontol. 2013;48(5):517-524.
doi pubmed - Go EK, Jung KJ, Kim JY, Yu BP, Chung HY. Betaine suppresses proinflammatory signaling during aging: the involvement of nuclear factor-kappaB via nuclear factor-inducing kinase/IkappaB kinase and mitogen-activated protein kinases. J Gerontol A Biol Sci Med Sci. 2005;60(10):1252-1264.
doi pubmed - Rehman A, Mehta KJ. Betaine in ameliorating alcohol-induced hepatic steatosis. Eur J Nutr. 2022;61(3):1167-1176.
doi pubmed pmc - Nemmar A, Yuvaraju P, Beegam S, Ali BH. Betaine (N,N,N-trimethylglycine) averts photochemically-induced thrombosis in pial microvessels in vivo and platelet aggregation in vitro. Exp Biol Med (Maywood). 2015;240(7):955-960.
doi pubmed pmc - Schwab U, Torronen A, Toppinen L, Alfthan G, Saarinen M, Aro A, Uusitupa M. Betaine supplementation decreases plasma homocysteine concentrations but does not affect body weight, body composition, or resting energy expenditure in human subjects. Am J Clin Nutr. 2002;76(5):961-967.
doi pubmed - Konstantinova SV, Tell GS, Vollset SE, Nygard O, Bleie O, Ueland PM. Divergent associations of plasma choline and betaine with components of metabolic syndrome in middle age and elderly men and women. J Nutr. 2008;138(5):914-920.
doi pubmed - Huang RZ, Ma JF, Chen S, Chen YM, Fang AP, Lu XT, Huang ZH, et al. Associations of serum betaine with blood pressure and hypertension incidence in middle-aged and older adults: a prospective cohort study. Food Funct. 2023;14(10):4881-4890.
doi pubmed - Rosas-Rodriguez JA, Sonanez-Organis JG, Godoy-Lugo JA, Espinoza-Salazar JA, Lopez-Jacobo CJ, Stephens-Camacho NA, Gonzalez-Ochoa G. Betaine Aldehyde Dehydrogenase expression during physiological cardiac hypertrophy induced by pregnancy. Biochem Biophys Res Commun. 2017;490(3):623-628.
doi pubmed - Steenge GR, Verhoef P, Katan MB. Betaine supplementation lowers plasma homocysteine in healthy men and women. J Nutr. 2003;133(5):1291-1295.
doi pubmed - Holm PI, Ueland PM, Vollset SE, Midttun O, Blom HJ, Keijzer MB, den Heijer M. Betaine and folate status as cooperative determinants of plasma homocysteine in humans. Arterioscler Thromb Vasc Biol. 2005;25(2):379-385.
doi pubmed - Ashtary-Larky D, Bagheri R, Ghanavati M, Asbaghi O, Tinsley GM, Mombaini D, Kooti W, et al. Effects of betaine supplementation on cardiovascular markers: A systematic review and Meta-analysis. Crit Rev Food Sci Nutr. 2022;62(23):6516-6533.
doi pubmed - Frey N, Katus HA, Olson EN, Hill JA. Hypertrophy of the heart: a new therapeutic target? Circulation. 2004;109(13):1580-1589.
doi pubmed - Giachini FR, Webb RC, Tostes RC. STIM and Orai proteins: players in sexual differences in hypertension-associated vascular dysfunction? Clin Sci (Lond). 2009;118(6):391-396.
doi pubmed pmc - Lacruz RS, Feske S. Diseases caused by mutations in ORAI1 and STIM1. Ann N Y Acad Sci. 2015;1356(1):45-79.
doi pubmed pmc - Luo R, Gomez AM, Benitah JP, Sabourin J. Targeting orai1-mediated store-operated Ca(2+) entry in heart failure. Front Cell Dev Biol. 2020;8:586109.
doi pubmed pmc
This article is distributed under the terms of the Creative Commons Attribution Non-Commercial 4.0 International License, which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Cardiology Research is published by Elmer Press Inc.